In this video, we're going to begin our discussion on the lock and key and the induced fit models. So it turns out that there are 2 main models for enzyme substrate specificity, or 2 main models for how the enzyme interacts with the substrate to form the enzyme substrate complex. And so the first main model is the lock and key model, whereas the second main model is the induced fit model. And so in this video, we're only going to talk about the lock and key model, but in our next lesson video, we'll talk about the induced fit model. And so with the lock and key model, the shape of the active site on the enzyme is rigid and perfectly complementary to the substrate's shape. Complementary to the substrate's shape. And so what this means is that in the lock and key model, the shape of the active site is perfectly suited to the shape of the substrate to create a perfect puzzle piece-like fit. And so if we take a look at our example down below of the lock and key model, notice that our enzyme is in purple here and our substrate is this key shape here. And so, in the lock and key model, the substrate acts as the key and the enzyme acts as the lock. And in the lock and key model, the active site of the enzyme takes on a shape that is perfectly complementary to the shape of the substrate. So when the enzyme substrate complex forms, over here, you can see that the substrate fits into the active site like a perfect puzzle piece-like fit. And so there's no need for the enzyme or the substrate to have to change conformations in order for the enzyme substrate complex to form. And so ultimately, what this means is that in the lock and key model, the enzyme substrate complex that forms is going to be very stable, almost way too stable in some cases. And so over the years, the lock and key model has actually become a less likely model for enzyme catalysis, and this has to do with the fact that the enzyme substrate complex becomes so stabilized with this lock and key model. And, ultimately, the stabilization of the enzyme substrate complex leads to the energy of activation either staying exactly the same, or even potentially increasing in comparison to the uncatalyzed reaction. And recall, that's the exact opposite effect that we want enzymes to have on reactions. We want enzymes to decrease the energy of activation, not to keep the energy of activation the same or even increase them, and that is again why the lock and key model is a less likely model. So, let's take a look down below at our energy diagram that we have of the lock and key model, where we have the free energy on the y-axis and the reaction coordinate on the x-axis. And notice that the dotted line curve right here represents the uncatalyzed reaction, whereas the red curve here represents the enzyme catalyzed reaction. And notice that this black line right here, this black arrow, represents the energy of activation for the uncatalyzed reaction and typically, enzymes are supposed to decrease the energy of activation for a reaction, but notice that the enzyme catalyzed activation energy represented by this red arrow here is pretty much exactly the same size as the uncatalyzed energy of activation. And that has to do with the fact that the lock and key model stabilizes the energy, the enzyme substrate complex, the energy of the enzyme substrate complex. So this low energy of the enzyme substrate complex ultimately leads to the energy of activation, this energy barrier represented by this red arrow here, being exactly the same size or becoming even larger depending on how well this transition state is stabilized. And so, ultimately, this is why the lock and key model is a less likely model for enzyme catalysis. And so a more likely model for enzyme catalysis is the induced fit model, which we'll talk about in our next lesson video. So I'll see you guys there.
Lock and Key Vs. Induced Fit Models - Online Tutor, Practice Problems & Exam Prep
 Created using AI
Created using AIThe lock and key model describes enzyme-substrate interaction as a rigid fit, where the enzyme's active site is perfectly complementary to the substrate's shape, leading to a stable enzyme-substrate complex. However, this model is less likely because it does not effectively lower the activation energy. In contrast, the induced fit model allows for conformational changes in both the enzyme and substrate, prioritizing stabilization of the transition state, which decreases the activation energy and enhances reaction rates, making it a more accurate representation of enzyme catalysis.
Lock-and-Key Vs. Induced Fit Models
Video transcript
Lock and Key Vs. Induced Fit Models
Video transcript
So in our last lesson video, we said that the lock and key model is a less likely model for enzyme substrate specificity. A more likely model for enzyme substrate specificity is the induced fit model, which is what we're going to talk about in this video. So with the induced fit model, unlike the lock and key model, the shape of the active site on the enzyme is not rigid. It's actually adjustable. Instead of being perfectly complementary to the shape of the substrate, the active site's shape is actually more complementary to the transition state than it is to the shape of the substrate. Ultimately, this allows the enzyme to prioritize stabilizing the transition state over stabilizing the enzyme-substrate complex, like what happens with the lock and key model.
As we'll see down below in our example, with the induced fit model, there are conformational changes that take place and are induced in both the enzyme's active site and the substrate. If we take a look at our example of the induced fit model down below, notice that we have the enzyme over here in purple and then we have our square-shaped substrate over here in orange. Notice that with the induced fit model, the active site shape is not perfectly complementary to the shape of the substrate, and instead, the active site shape is more complementary to the transition state. When the substrate forms a complex with the enzyme, notice that there are conformational changes that are induced in both the enzyme's active site and the substrate.
So notice that the square-shaped substrate has changed conformations to this rectangular-shaped substrate. Also, notice that the enzyme's active site is more opened up and has essentially changed conformations. Ultimately, what happens when the enzyme-substrate complex forms in the induced fit model is that the enzyme-substrate complex is not stabilized like it was with the lock and key model. Instead, ultimately, what this means is that the transition state stabilization is prioritized with the induced fit model, and that is ultimately what makes the induced fit model a more likely model for enzyme catalysis. This again has to do with the fact that the enzyme-substrate complex is not stabilized like it is with the lock and key model. And when the transition state is stabilized and prioritized, ultimately what happens is there's a decrease in the energy of activation. And that's exactly what we want enzymes to do, and that's why the induced fit model is more likely.
If we take a look down below at our energy—this dotted line here that represents our uncatalyzed reaction, whereas the red line represents the enzyme-catalyzed reaction. And notice that the arrow here, the black arrow, represents the energy of activation for the uncatalyzed reaction, which is pretty large. And notice that this time the red arrow is representing the catalyzed energy of activation and it's actually smaller than the uncatalyzed energy of activation, which means that the reaction is going to be sped up. So the reason that we're able to get a smaller energy of activation for the catalyzed reaction is that notice that the enzyme-substrate complex is not as stabilized as it was with the lock and key model, and that allows for us to just stabilize the transition state and keep the energy of activation for the catalyzed reaction relatively small, instead of staying the same or potentially increasing, like it did with the lock and key model.
It's important to note, moving forward in our course, that the induced fit model is a more likely model for enzyme catalysis. That concludes our lesson on the induced fit model, and I'll see you guys in our next video where we'll be able to get a little bit of practice.
Compare the two enzyme-catalyzed reaction diagrams below (i & ii) to determine which of the following is true.
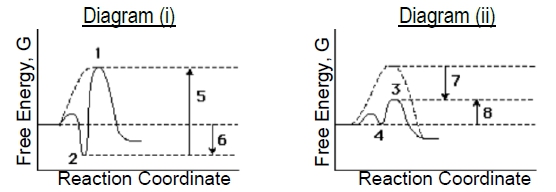
a) The ES-complex in diagram (i) is #2 and in diagram (ii) is #3..
b) Catalyzed EA in diagram (i) is arrow #5 and in diagram (ii) is arrow #7.
c) Binding energy in diagram (i) is arrow #5 and in diagram (ii) is arrow #7.
d) Diagram (i) describes a 'lock & key' model while (ii) describes more of an 'induced fit' model.
What is a potential disadvantage for an enzyme having too high of an affinity for its substrate?
Select the best option that fills in the blanks appropriately in the order of their appearance.
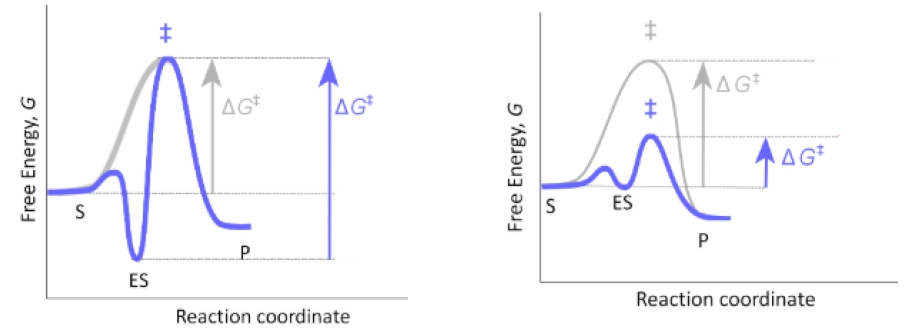
The left graph depicts an energy diagram for the '_____________' model of enzyme-substrate specificity, whereas the right diagram depicts an energy diagram for the modern '_____________' model. In the lock & key model, the enzyme binds tightly and precisely to the '_____________.' In the induced fit model, the enzyme binds weakly to the '_____________' and then changes conformation to bind tightly to the '_____________.'
Here’s what students ask on this topic:
What is the lock and key model in enzyme-substrate interaction?
The lock and key model describes enzyme-substrate interaction as a rigid fit, where the enzyme's active site is perfectly complementary to the substrate's shape. This model likens the enzyme to a lock and the substrate to a key, suggesting that only the correctly shaped substrate (key) can fit into the enzyme's active site (lock). This results in a stable enzyme-substrate complex. However, this model is less favored because it does not effectively lower the activation energy required for the reaction, which is a key function of enzymes.
Created using AIWhy is the lock and key model considered less likely for enzyme catalysis?
The lock and key model is considered less likely for enzyme catalysis because it leads to an overly stable enzyme-substrate complex. This excessive stabilization does not lower the activation energy of the reaction, which is contrary to the primary role of enzymes. Enzymes are supposed to decrease the activation energy to speed up reactions. In the lock and key model, the activation energy remains the same or may even increase, making it an inefficient model for explaining enzyme activity.
Created using AIWhat is the induced fit model in enzyme-substrate interaction?
The induced fit model describes enzyme-substrate interaction as a dynamic process where the enzyme's active site is flexible and adjusts its shape to fit the substrate. Unlike the lock and key model, the active site is more complementary to the transition state than to the substrate itself. This flexibility allows for conformational changes in both the enzyme and the substrate, leading to a less stable enzyme-substrate complex but a more stabilized transition state. This stabilization of the transition state effectively lowers the activation energy, making the induced fit model a more accurate representation of enzyme catalysis.
Created using AIHow does the induced fit model lower the activation energy of a reaction?
The induced fit model lowers the activation energy of a reaction by prioritizing the stabilization of the transition state rather than the enzyme-substrate complex. When the substrate binds to the enzyme, both undergo conformational changes that make the active site more complementary to the transition state. This stabilization of the transition state reduces the energy barrier (activation energy) required for the reaction to proceed, thereby speeding up the reaction. This is in contrast to the lock and key model, where the enzyme-substrate complex is overly stabilized, failing to lower the activation energy effectively.
Created using AIWhat are the main differences between the lock and key and induced fit models?
The main differences between the lock and key and induced fit models lie in the flexibility and stabilization of the enzyme-substrate complex. In the lock and key model, the enzyme's active site is rigid and perfectly complementary to the substrate, leading to a highly stable enzyme-substrate complex but failing to lower the activation energy. In contrast, the induced fit model features a flexible active site that adjusts to fit the substrate, prioritizing the stabilization of the transition state. This flexibility and transition state stabilization effectively lower the activation energy, making the induced fit model a more accurate representation of enzyme catalysis.
Created using AI