So now that we've differentiated between passive and active membrane transport, in this video we're going to talk more details about passive membrane transport. And so if we take a look at our membrane transport map over here, what you'll notice is that passive transport can be further categorized into simple passive transport or simple diffusion, and facilitated passive transport or facilitated diffusion. And really that's the main focus of this video that there are 2 types of passive transport. Simple and facilitated passive transport. And so the first type the simple passive transport or simple diffusion is as simple as it sounds. Simple diffusion is going to be non energetic, simple, and direct diffusion straight through the membrane, And this means that the molecules are going to be squeezing between the phospholipids in order to diffuse from an area of high concentration to an area of low concentration. Here, the facilitated passive transport or facilitated diffusion, again is also going to be non energetic because it is passive, and we know passive means no energy. And so this is non energetic diffusion that is facilitated by a membrane transport protein. However, again, the membrane transport protein does not utilize ATP or energy since it's non energetic. And so down below in our example, here, we can differentiate between the 2 different types of passive transport. Simple passive transport versus facilitated Passive Transport or diffusion. And so, over here on the left hand side of this image what we're showing you is simple diffusion, and really what you can see is that the molecules are gonna be transported across a biological membrane from an area of high concentration to an area of low concentration, but because it's simple it does not require facilitation from a protein. So these molecules are able to squeeze their way between the phospholipids just as we mentioned up above to get to the other side of the membrane. And so molecules that are capable of simple diffusion are going to be very very small and very non polar molecules, such as the gases carbon dioxide, oxygen, and nitrogen gas. Now, on the other hand over here on this side of our image, what we have is facilitated passive transport or facilitated diffusion, and so notice that the molecules are still being transported from an area of high concentration down to an area of low concentration. So still no energy is needed. This is still non energetic. However, what you'll notice is different is that there is this membrane protein here that is required to facilitate the diffusion. And so what you'll notice here is that we have molecules here that are charged, and we know that charged molecules can't really cross a membrane without facilitation. However, if they do have facilitation, they are able to be transported passively down their concentration gradient just as we see here, and that is what we're calling facilitated diffusion. And so this here concludes our video distinguishing the 2 different types of passive membrane transport, which are simple and facilitated passive transport. And so we'll be able to get some practice applying these concepts in our next few videos. So I'll see you guys there.
Passive Membrane Transport: Study with Video Lessons, Practice Problems & Examples
 Created using AI
Created using AIPassive membrane transport consists of two types: simple diffusion and facilitated diffusion. Simple diffusion occurs without energy, allowing small, nonpolar molecules like oxygen and carbon dioxide to pass directly through the lipid bilayer. In contrast, facilitated diffusion also requires no energy but utilizes membrane transport proteins for charged molecules. The kinetics of these processes differ; simple diffusion shows a linear relationship with concentration, while facilitated diffusion forms a hyperbolic curve due to limitations imposed by the number of transport proteins, akin to enzyme kinetics.
Passive Membrane Transport
Video transcript
The difference between simple and facilitated diffusion is that facilitated diffusion:
Simple diffusion and facilitated diffusion across a membrane both ___________________:
Passive Membrane Transport
Video transcript
So now that we've differentiated between the 2 different types of passive membrane transport, simple diffusion and facilitated diffusion, in this video, we're going to talk a little bit about the kinetics of passive transport, which is going to help us even further distinguish between simple diffusion and facilitated diffusion as we'll see. Now you might recall that way back in our previous lesson videos, we actually covered enzyme kinetics. And from those old videos on enzyme kinetics, we already know that kinetics is just the field that studies the rates, velocities, and speeds of particular processes. And so the rate or velocity of passive transport is driven by the extent of the concentration gradient across the membrane. And so the greater the concentration is of the diffusing substance on one side of the membrane, the faster the rate of simple diffusion. And so of course what this means is that the rate of simple diffusion will form linear data when plotted onto a graph like what we have down below right over here. And so this graph should look familiar to you guys because it's the same exact graph from our old lesson videos on enzyme kinetics. And so on the y-axis, what we have is \( v_0 \), which is the initial reaction velocity. And of course, the reaction here in this scenario is just diffusion, and so \( v_0 \) is just the initial velocity of diffusion. And on the x-axis, what we have is the substrate concentration in quotes because technically it's not a substrate if diffusion is the only thing that's occurring. But really you can just think of the substrate concentration in quotes here as just the concentration of the diffusing substance. And so right now we're just focusing on this green line right here for simple diffusion, and the most important thing that you should take away here from this green line is that simple diffusion will form linear data when plotted onto this graph. And so as we increase the concentration of the diffusing substance from left to right, we will also increase the rate or the velocity of simple diffusion in the same manner and that's what creates this linear data that we see right here. And that's exactly what we said up above right here. The greater the concentration, the faster the rate of simple diffusion, forming linear data. And so, of course, because linear data is associated with simple diffusion, this means that the data is explained by the equation of a line which recall is just \( y = mx + b \), and so so far this is pretty easy pretty straightforward. Simple diffusion forms a linear straight line. Now in terms of the rate of facilitated diffusion, on the other hand, you may have already noticed that it's a little bit different by looking at this blue curve that we have here. And so one of the first differences you probably noticed is that simple diffusion forms a straight linear line whereas facilitated diffusion forms this curve, But another thing that you should notice is that the rate of facilitated diffusion is even faster than the rate of simple diffusion at any concentration of diffusing substance. And so no matter what value of concentration of diffusing substance you choose here, if you go up and look at the velocity levels that correspond with them, what you'll notice is that the rate of facilitated diffusion is always faster than the rate of simple diffusion. Now, although the rate of facilitated diffusion is even faster than the rate of simple diffusion, there is one limitation that we should take into account, however, and that is that the rate of facilitated diffusion is limited, and it's limited by the amount of transport protein that's present in the membrane. And so what this means is that ultimately when we increase this, the concentration of diffusing substance enough, the rate of facilitated diffusion will reach a limitation, a \( V_{\text{max}} \) if you will, a maximum rate of diffusion. And this maximum rate of diffusion is again limited by the amount of transport protein. And so this limitation here is very similar to how an enzyme catalyzed reaction is limited by the amount of enzyme. And so you can really think of the transport protein that's involved in facilitated diffusion as being very similar to an enzyme in that respect in terms of the limitation. And so because facilitated diffusion is limited by the amount of transport protein, therefore what this means is that passive facilitated diffusion rates, instead of forming linear data, they are going to form a hyperbolic Michaelis Menten type curve. And so that's exactly why we have this curve right here for facilitated diffusion approaching some kind of maximum velocity that's limiting it, and again that's limited by the amount of transport protein. And so the biggest takeaway here is that facilitated diffusion is going to be hyperbolic, form hyperbolic data, whereas simple diffusion will form linear data. And so, of course, this hyperbolic curve right here, the data is explained through the Michaelis Menten equation, which is what we have right here. And so, really the only difference here is that we have \( K_{\text{TR}} \) instead of the Michaelis constant \( K_m \) that we're used to seeing in our enzyme kinetics. But really the \( K_{\text{TR}} \) is pretty much the same exact thing. It's analogous to the Kilometers, and so the \( K_{\text{TR}} \) would represent the substrate concentration at which the transport protein is half saturated. And so you can see that the \( t_{\text{r}} \) in transport reminds us that it's going to be for the diffusion and the transport of a specific molecule. And so, again, the biggest takeaways that you guys should get from this video is that simply by plotting the data onto a graph like this, a biochemist can tell if a molecule is diffusing via facilitation because it will form a hyperbolic Michaelis Menten curve, or if the molecule is diffusing by simple diffusion because it forms linear data that's explained by the equation of a line. And so, that will help us differentiate between facilitated and simple diffusion even more. And that concludes our lesson on the kinetics of passive transport and we'll be able to get some practice applying these concepts in our next couple of videos. So I'll see you guys there.
The rate of movement (flux) of a substance X into cells was measured at different concentrations of X to construct the following graph. Does the graph's data suggest diffusion of X into cells is mediated by a protein transporter?
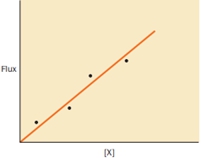
The rate of transfer across a membrane is measured for a given molecule. The diffusion rate is seen to be hyperbolic with respect to the concentration of the diffusing molecule. The method of transport is which of the following?
Here’s what students ask on this topic:
What is the difference between simple diffusion and facilitated diffusion in passive membrane transport?
Simple diffusion and facilitated diffusion are both types of passive membrane transport, meaning they do not require energy. Simple diffusion allows small, nonpolar molecules like oxygen and carbon dioxide to pass directly through the lipid bilayer by squeezing between phospholipids. In contrast, facilitated diffusion involves the use of membrane transport proteins to help charged or larger molecules cross the membrane. While both processes move substances from an area of high concentration to low concentration, facilitated diffusion is limited by the number of available transport proteins, leading to a hyperbolic relationship between concentration and rate, unlike the linear relationship seen in simple diffusion.
Created using AIHow does the concentration gradient affect the rate of simple diffusion?
The rate of simple diffusion is directly influenced by the concentration gradient across the membrane. The greater the concentration difference of the diffusing substance on either side of the membrane, the faster the rate of diffusion. This relationship is linear, meaning that as the concentration of the diffusing substance increases, the rate of diffusion increases proportionally. This can be represented graphically, where the initial velocity of diffusion (v0) plotted against the concentration of the diffusing substance forms a straight line, described by the equation of a line, y = mx + b.
Created using AIWhy does facilitated diffusion show a hyperbolic curve in its kinetics?
Facilitated diffusion shows a hyperbolic curve in its kinetics because it is limited by the number of transport proteins available in the membrane. As the concentration of the diffusing substance increases, the rate of facilitated diffusion initially increases rapidly. However, once all transport proteins are saturated, the rate of diffusion reaches a maximum (Vmax), creating a plateau in the curve. This behavior is similar to enzyme kinetics and can be described by the Michaelis-Menten equation, where the transport rate is a function of the concentration of the diffusing substance and the number of available transport proteins.
Created using AIWhat types of molecules typically use simple diffusion to cross the cell membrane?
Molecules that typically use simple diffusion to cross the cell membrane are small and nonpolar. These include gases like oxygen (O2), carbon dioxide (CO2), and nitrogen (N2). Because these molecules are nonpolar, they can easily pass through the hydrophobic core of the lipid bilayer without the need for transport proteins. Simple diffusion allows these molecules to move from an area of high concentration to an area of low concentration directly through the membrane.
Created using AIHow can a biochemist determine if a molecule is diffusing via simple or facilitated diffusion?
A biochemist can determine if a molecule is diffusing via simple or facilitated diffusion by plotting the rate of diffusion against the concentration of the diffusing substance. If the data forms a linear relationship, it indicates simple diffusion. However, if the data forms a hyperbolic curve, it suggests facilitated diffusion. This hyperbolic curve is due to the saturation of transport proteins, similar to the Michaelis-Menten kinetics observed in enzyme-catalyzed reactions. The linear data for simple diffusion can be described by the equation of a line (y = mx + b), while the hyperbolic data for facilitated diffusion follows the Michaelis-Menten equation.
Created using AI