Back
BackProblem 1
Explain why many developmental genes encode either transcription factors or signaling molecules.
Problem 2
Bird beaks develop from an embryonic group of cells called neural crest cells that are part of the neural tube, which gives rise to the spinal column and related structures. Amazingly, neural crest cells can be surgically transplanted from one embryo to another, even between embryos of different species. When quail neural crest cells were transplanted into duck embryos, the beak of the host embryo developed into a shape similar to that found in quails, creating the 'quck.' Duck cells were recruited in addition to the quail cells to form part of the quck beak. Conversely, when duck neural crest cells were transplanted into quail embryos, the beak of the embryo resembled that of a duck, creating a 'duail,' and quail cells were recruited to form part of the beak. What do these experiments tell you about the autonomy or nonautonomy of the transplanted and host cells during beak development?
Problem 3
How is positional information provided along the anterior–posterior axis in Drosophila? What are the functions of bicoid and nanos?
Problem 4a
Early development in Drosophila is atypical in that pattern formation takes place in a syncytial blastoderm, allowing free diffusion of transcription factors between nuclei. In many other animal species, the fertilized egg is divided by cellular cleavages into a larger and larger number of smaller and smaller cells.
What constraints does the formation of a syncytial blastoderm impose on the mechanisms of pattern formation?
Problem 4b
Early development in Drosophila is atypical in that pattern formation takes place in a syncytial blastoderm, allowing free diffusion of transcription factors between nuclei. In many other animal species, the fertilized egg is divided by cellular cleavages into a larger and larger number of smaller and smaller cells.
How must the model that describes Drosophila development be modified for describing animal species whose early development is not syncytial?
Problem 5a
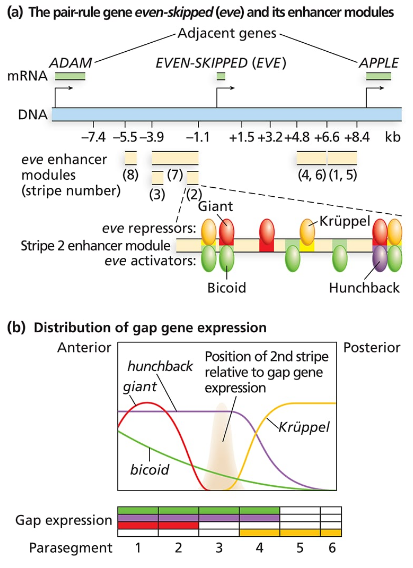
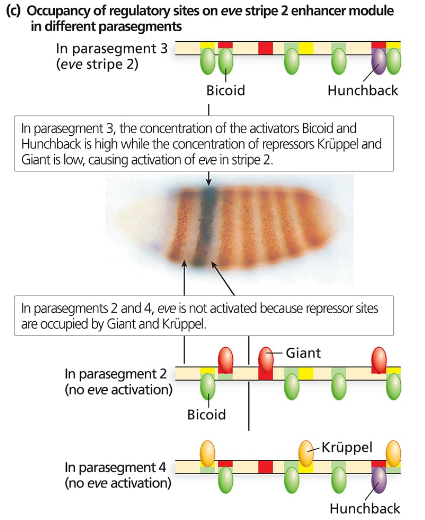
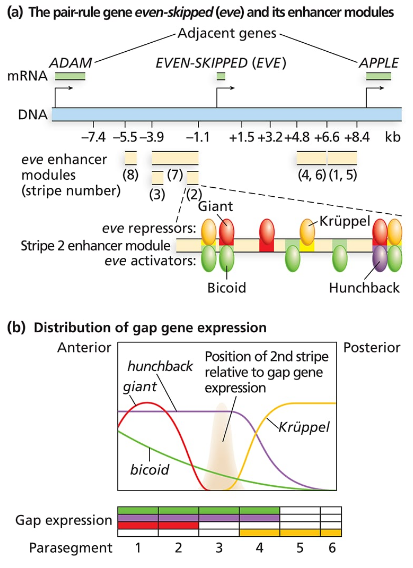
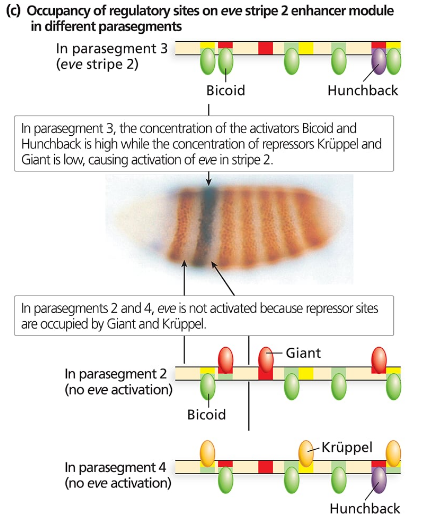
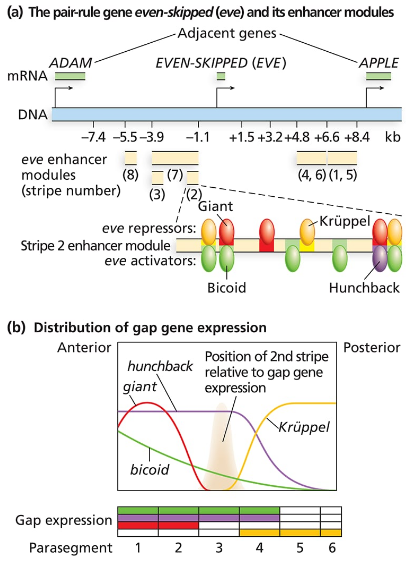
Consider the even-skipped regulatory sequences in the following figure:
How are the sharp boundaries of expression of Eve Stripe 2 formed?
Problem 5b
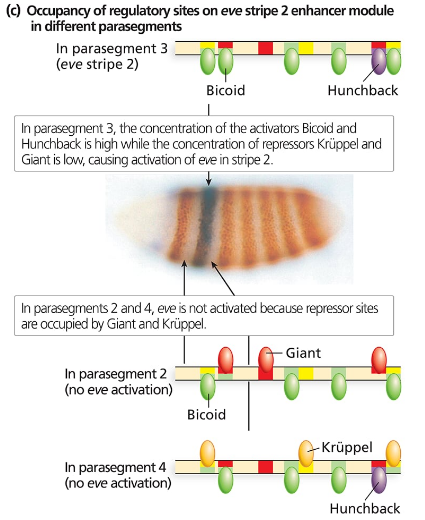
Consider the even-skipped regulatory sequences in Figure 18.9.
Consider the binding sites for gap proteins and Bicoid in the stripe 2 enhancer module. What sites are occupied in parasegments 2, 3, and 4, and how does this result in expression or no expression?
Problem 5c
Consider the even-skipped regulatory sequences in Figure 18.9.
Explain what you expect to see happen to even-skipped stripe 2 if it is expressed in a Krüppel mutant background. What about a hunchback mutant background? A giant mutant background? A bicoid mutant background?
Problem 6
What is the difference between a parasegment and a segment in Drosophila development? Why do developmental biologists think of parasegments as the subdivisions that are produced during the development of flies?
Problem 7
Why do loss-of-function mutations in Hox genes usually result in embryo lethality, whereas gain-of-function mutants can be viable? Why are flies homozygous for the recessive loss-of-function alleles and viable?
Problem 8
Compare and contrast the specification of segmental identity in Drosophila with that of floral organ specification in Arabidopsis. What is the same in this process, and what is different?
Problem 9
Actinomycin D is a drug that inhibits the activity of RNA polymerase II. In the presence of actinomycin D, early development in many vertebrate species, such as frogs, can proceed past the formation of a blastula, a hollow ball of cells that forms after early cleavage divisions, but development ceases before gastrulation (the stage at which cell layers are established). What does this tell you about maternal versus zygotic gene activity in early frog development?
Problem 10a
Ablation of the anchor cell in wild-type C. elegans results in a vulva-less phenotype.
What phenotype is to be expected if the anchor cell is ablated in a let-23 loss-of-function mutant?
Problem 10b
Ablation of the anchor cell in wild-type C. elegans results in a vulva-less phenotype.
What about if the anchor cell is ablated in a let-23 gain-of-function mutant?
Problem 11
In gain-of-function let-23 and let-60 C. elegans mutants, all of the vulval precursor cells differentiate with 1° or 2° fates. Do you expect adjacent cells to differentiate with 1° fates or with 2° fates? Explain.
Problem 12a
In mammals, identical twins arise when an embryo derived from a single fertilized egg splits into two independent embryos, producing two genetically identical individuals.
What limits might there be, from a developmental genetic viewpoint, as to when this can occur?
Problem 12b
In mammals, identical twins arise when an embryo derived from a single fertilized egg splits into two independent embryos, producing two genetically identical individuals.
The converse phenotype, fusion of two genetically distinct embryos into a single individual, is also known. What are the genetic implications of such an event?
Problem 13a
The bicoid gene is a coordinate maternal-effect gene.
A female Drosophila heterozygous for a loss-of-function bicoid allele is mated to a male that is heterozygous for the same allele. What are the phenotypes of their progeny?
Problem 13b
The bicoid gene is a coordinate maternal–effect gene. A female that is homozygous for a loss-of-function bicoid allele is mated to a wild-type male. What are the phenotypes of their progeny?
Problem 13c
The bicoid gene is a coordinate maternal–effect gene. If loss of bicoid function in the egg leads to lethality during embryogenesis, how are females homozygous for bicoid produced? What is the phenotype of a male homozygous for bicoid loss-of-function alleles?
Problem 14
Given that maternal Bicoid activates the expression of hunchback, what would be the consequence of adding extra copies of the bicoid gene by transgenic means to a wild-type female with two copies, thus creating a female fly with three or four copies of the bicoid gene? How would the hunchback expression be altered? What about the expression of other gap genes and pair-rule genes?
- What phenotypes do you expect in flies homozygous for loss-of-function mutations in the following genes: Krüppel, odd-skipped, hedgehog, and Ultrabithorax?
Problem 15
Problem 16a
The pair-rule gene fushi tarazu is expressed in the seven even-numbered parasegments during Drosophila embryogenesis. In contrast, the segment polarity gene engrailed is expressed in the anterior part of each of the 14 parasegments. Since both genes are active at similar times and places during development, it is possible that the expression of one gene is required for the expression of the other. This can be tested by examining expression of the genes in a mutant background—for example, looking at fushi tarazu expression in an engrailed mutant background, and vice versa. Given the hierarchy of gene action during Drosophila embryogenesis, what might you predict to be the result of these experiments?
Problem 16b
The pair-rule gene fushi tarazu is expressed in the seven even-numbered parasegments during Drosophila embryogenesis. In contrast, the segment polarity gene engrailed is expressed in the anterior part of each of the 14 parasegments. Since both genes are active at similar times and places during development, it is possible that the expression of one gene is required for the expression of the other. This can be tested by examining the expression of the genes in a mutant background—for example, looking at fushi tarazu expression in an engrailed mutant background, and vice versa. Based on your prediction, can you predict the phenotype of the fushi tarazu and engrailed double mutant?
Problem 17
In contrast to Drosophila, some insects (e.g., centipedes) have legs on almost every segment posterior to the head. Based on your knowledge of Drosophila, propose a genetic explanation for this phenotype, and describe the expected expression patterns of genes of the Antennapedia and bithorax complexes.
- The bristles that develop from the epidermis in Drosophila are evenly spaced, so that two bristles never occur immediately adjacent to each other. How might this pattern be established during development?
Problem 18
Problem 19
You are traveling in the Netherlands and overhear a tulip breeder describe a puzzling event. Tulips normally have two outer whorls of brightly colored petal-like organs, a third whorl of stamens, and an inner (fourth) whorl of carpels. However, the breeder found a recessive mutant in his field in which the outer two whorls were green and sepal-like, whereas the third and fourth whorls both contained carpels. What can you speculate about the nature of the gene that was mutated?
Problem 20a
A powerful approach to identifying genes of a developmental pathway is to screen for mutations that suppress or enhance the phenotype of interest. This approach was undertaken to elucidate the genetic pathway controlling C. elegans vulval development. A lin-3 loss-of-function mutant with a vulva-less phenotype was mutagenized. Based on your knowledge of the genetic pathway, what types of mutations will suppress the vulva-less phenotype?
Problem 20b
A powerful approach to identifying genes of a developmental pathway is to screen for mutations that suppress or enhance the phenotype of interest. This approach was undertaken to elucidate the genetic pathway controlling C. elegans vulval development. In a complementary experiment, a gain-of-function let-23 mutant with a multi-vulva phenotype was also mutagenized. What types of mutations will suppress the multi-vulva phenotype?
Problem 21a
The Hoxd9–13 genes are thought to specify digit identity. What would be the consequence of ectopically expressing Hoxd10 throughout the developing mouse limb bud? What about Hoxd11? What about both Hoxd10 and Hoxd11?