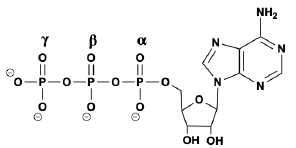
Phosphorylation is a crucial type of post-translational modification involving the covalent attachment of a phosphate group to a protein or enzyme. This process plays a significant role in enzyme regulation, making it one of the most important mechanisms for controlling protein activity. The addition of a phosphate group can either activate or inactivate a protein, depending on the context. This dual functionality is essential for regulating various biological processes.
When a phosphate group is added to a protein, it can lead to a change in the protein's conformation, which is the three-dimensional shape that determines its function. For instance, a protein may exist in a circular form before phosphorylation and adopt a different conformation afterward, which can enhance or inhibit its activity. Understanding phosphorylation is vital for comprehending how enzymes are regulated and how cellular processes are controlled.
In summary, phosphorylation is a key regulatory mechanism that alters protein function through the addition of phosphate groups, impacting the protein's shape and activity. This modification is fundamental to many cellular processes, and further exploration of the sources of phosphate groups will enhance our understanding of this important biochemical phenomenon.