Alright. So here we're going to revisit our map of the lesson on biosignalling pathways which we have down below. And of course, we know that we're exploring this map by exploring the leftmost branches first, and so we've already talked about G protein-coupled receptors or GPCRs, and we've talked specifically about the stimulatory adenylate cyclase GPCR signaling pathway, and we've talked in detail about the secondary messenger cAMP and the enzyme PKA in this pathway. And so now that we've explored this pathway here to its full extent, we're now going to shift over and talk about this pathway over here. And so this is going to be the inhibitory pathway for adenylate cyclase GPCR signaling. So we're still talking about the same pathway except the inhibitory pathway which is going to have some differences. And so, let's go on and get started talking about this pathway. So here we're going to introduce inhibitory adenylate cyclase GPCR signaling, which is not to be confused with what we covered in our previous lesson videos, which was stimulatory not inhibitory adenylate cyclase GPCR signaling. And so what we need to recall from our previous lesson videos is that not all G proteins stimulate or activate the effector enzyme. In fact, there are some G proteins that do the complete opposite and inhibit the effector enzyme. Now this Inhibitory Adenylate Cyclase GPCR signaling has the same exact pathway as before, except this time we have an inhibitory alpha subunit of the G protein or a GI that ultimately inhibits the activity of the effector enzyme, so that it creates less secondary messenger than before, and we'll be able to see this down below in our image. Now the integration of what we covered in our previous lesson videos, which was the stimulatory pathway, and what we're talking about now, which is the inhibitory GPCR pathway, these 2 can be integrated together and can help to regulate the activity of the effector enzyme so that the effector enzyme is creating the exact amount, the appropriate amount of secondary messenger that the cell needs. And so if we take a look at our image down below, at our example image, notice that we're showing you the activity of the effector enzyme Adenylate Cyclase being regulated by the inhibitory GPCR pathway. And so on the right side of our image over here, notice we're showing you stimulation or the stimulatory pathway utilizing the stimulatory G protein GS, And on the right hand side of the image over here, we're showing you the inhibitory G protein pathway or GI. And so on the left hand side, it's pretty much exactly what we talked about in our previous lesson video. So you can see epinephrine binds to the beta-adrenergic GPCR causing a conformational shift that activates the G protein, causes it to swap out its GDP and replace it with GTP, and then the alpha subunit of the G protein is going to dissociate and activate the adenylate cyclase effector enzyme so that it creates more, it converts more ATP into secondary messenger cAMP. And so really nothing new over here on the left-hand side of the image. Whereas if we take a look at the right-hand side of the image, this is really the new part that we're introducing here. And so there's some inhibitory ligand that we're not going to get into the details of that is going to bind to some inhibitory GPCR, that is going to induce a conformational shift in this inhibitory G PCR, which is going to activate an inhibitory G protein, by promoting again the GDP to be replaced with GTP just the same as it was over here. And so we have our dissociated GI, our dissociated, inhibitory G protein alpha subunit. And the only real difference between this side over here and the stimulatory side is what we see right here and over here. And so on the left-hand side, notice that the stimulatory pathway activates the adenylate cyclase, whereas the inhibitory pathway over here is going to inhibit adenylate cyclase so that it converts less ATP into cAMP. And so, really this inhibitory pathway over here is acting like the brakes in a car to help slow down the activity of adenylate cyclase. And really, this stimulatory pathway over here is almost acting like the gas pedal of a car to help speed up the activity of adenylate cyclase. And so you can see how the gas pedal and the brakes can be used to regulate the speed or the activity of the adenylate cyclase effector enzyme. And so really this here concludes our introduction to inhibitory adenylate cyclase GPCR signaling, and we'll be able to get some practice applying the concepts that we've learned here as we move forward in our course. So I'll see you guys in our next video.
Inhibitory Adenylate Cyclase GPCR Signaling - Online Tutor, Practice Problems & Exam Prep
 Created using AI
Created using AIBiosignaling pathways involve G protein-coupled receptors (GPCRs) that can either stimulate or inhibit adenylate cyclase activity. The stimulatory pathway increases cAMP production, while the inhibitory pathway, mediated by the inhibitory G protein (GI), decreases cAMP levels. Additionally, GPCR desensitization occurs when continuous ligand exposure dampens cellular response, primarily through the action of beta adrenergic GPCR kinase (BARK) and beta arrestin, which phosphorylate and block GPCR interactions, respectively. This regulation ensures appropriate cellular responses to signals.
Inhibitory Adenylate Cyclase GPCR Signaling
Video transcript
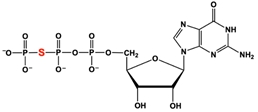
The addition of the nonhydrolyzable GTP-analog, GTPγS (shown below), is a common cell culture technique. If only affecting the inhibitory pathway, what effect would GTPγS have on cellular cAMP levels?
If a chemical is an inhibitor of the enzyme adenylyl cyclase, which of the following steps in the GPCR signaling pathway would be directly blocked?
Inhibitory Adenylate Cyclase GPCR Signaling
Video transcript
So here we're going to introduce yet another way to inhibit adenylate cyclase GPCR signaling, and that is through GPCR desensitization. Now when a signal is continuously present creating repeated exposure, then cells need the ability to desensitize to that continuously present signal that's again creating repeated exposure. Now the process of desensitization is really defined as the dampening or the decreasing of the cellular response even when the primary messenger or ligand still persists. And again, that primary ligand is continuously present creating repeated exposure. Now desensitization is really understandable when you think about it in the context of you getting up in the early morning and walking into a room with really bright lights. Initially, your eyes are going to create a really strong cell response to those bright lights and your eyes are going to squint when you walk into that room with bright lights. However, over time even when those bright lights still persist and those, that bright light signal is continuously present creating repeated exposure. Eventually, over time, your eyes will desensitize to those bright lights And so they will not create such a drastic cell response and eventually your eyes will be able to open back up to the normal position, that you have throughout the day. And so, the desensitization of your eyes to bright lights is really similar to how GPCRs can also desensitize to signals such as ligands that are continuously present.
And so really one of the major proteins that are involved in GPCR desensitization is the beta-adrenergic GPCR kinase, which as you can see by these bolded letters right here, is commonly referred to as just the BARK protein. And so, the beta-adrenergic GPCR kinase is indeed a kinase, so we know that it's going to phosphorylate things. And more specifically, it's going to phosphorylate the beta-adrenergic GPCR, which is the specific GPCR found in adenylate cyclase GPCR signaling. And so the BARK protein, because again it is a kinase, it's going to phosphorylate its substrate, and, again, its substrate is going to be C-terminal serine residues on the ligand-bound beta-adrenergic GPCR. And so ligand-bound is really important here because, again that is what's going to indicate that the signal is continuously present creating repeated exposure.
And so if we take a look down below at our image, over here on the left-hand side, notice that the very first step in GPCR desensitization is that the ligand that is continuously present creating repeated exposure, must actually bind to the beta-adrenergic GPCR. And, of course, we can indicate that the epinephrine, extracellular ligand must bind to the beta-adrenergic GPCR. And again, that is the very first step of GPCR desensitization. But then notice, moving on to the second step over here in our second image, notice that the beta-adrenergic GPCR kinase, or BARK, is coming into play, which is this yellow protein that we see down below. And so, again, because BARK is a kinase, it's going to phosphorylate the beta-adrenergic GPCR, as we see here, the phosphorylation. And so in step number 2 down below, we can indicate that BARK, this kinase, is going to again phosphorylate the beta-adrenergic GPCR.
Now another major protein that's involved in GPCR desensitization is the protein beta-arrestin, which you can see by these bolded letters right here is referred to as the BAR protein. And so the beta-arrestin or the BAR protein is again a protein that's going to bind to the phosphorylated GPCR, essentially blocking or preventing the GPCR's interaction with the G Protein. And ultimately, that's going to prevent the cell response from occurring and help to desensitize the GPCR from the signal. And so this beta-arrestin or BAR protein can actually initiate temporary endocytosis of the GPCR. And recall that endocytosis, the e n here, reminds us that it's going to allow to enter the cell as a, within a vesicle. And so this is going to make the GPCR temporarily inaccessible to the ligand that is continuously present, and that also helps to desensitize the GPCR, from the continuously present ligand.
And so if we take a look down below at our image here at our third step, notice that the beta-arrestin protein or the BAR protein is here in purple, and it is binding to the phosphorylated beta-adrenergic GPCR. And so beta-arrestin binds to that, phosphorylated GPCR, and essentially blocks it blocks its interaction with the g protein. So beta-arrestin blocks the g protein association with the gPCR, and it can also trigger temporary endocytosis of the GPCR, which we're not showing here, but you can imagine this entire thing being brought down into the cell via endocytosis, removing it from the plasma membrane so that it will not again cause a cell response. Now notice over here on the far right, we have this little memory tool over here to help you guys remember the roles of the BARK protein and the beta-arrestin protein in GPCR desensitization. And so you can imagine that GPCR desensitization, is only going to occur when the GPCR is kind of going rogue because there's so much signal that's present and the GPCR is just creating too much of a cell response.
And so here what we have is the GPCR kinda going rogue here. He's just doing whatever he wants and he's robbing a bank here as you can see. And so when you have a GPCR that's going rogue, you need a way to desensitize so that it does not you know, cause any issues. And so what can happen is the beta-adrenergic GPCR kinase or BARK here, like this dog barking, can essentially mark the rogue GPCR, and that is going to essentially allow the officer beta-arrestin to come in and arrest the GPCR and prevent it from interacting with the g protein money bag over here. So that way the GPCR is not going rogue and it's arrested temporarily until the signal kinda dampens down.
But this here concludes our introduction to GPCR desensitization, and we'll be able to get some practice with these concepts as we move forward in our course. So I'll see you guys in our next video.
Inhibitory Adenylate Cyclase GPCR Signaling Example 1
Video transcript
Alright. So here we have a word problem as an example, and it says that a cell line expressing the beta adrenergic GPCR is incubated in epinephrine for 5 minutes. The cells are then lysed or burst open, and the GPCR protein is purified. To determine if beta arrestin is bound to that purified GPCR, the purified GPCR solution is examined by Western blot using an antibody against beta arrestin, and that is really the data that we see in lanes number 1 and lanes number 2 down below in our image. And so taking a look at this image down below briefly, notice that we have the direction of the gel migration, so the top of the gel is the starting place and the bottom of the gel is the ending place. And then notice that we have the exposure to epinephrine indicated down below in terms of the time. So 0 minutes was in lane number 1, and 5 minutes was in lane number 2. Now notice that the problem goes on to say that the experiment was repeated, but this time prior to and during epinephrine addition, the cells are incubated in an inhibitor that blocks beta adrenergic GPCR phosphorylation by the GPCR Kinase BARK, and that is really the data that we see in lanes number 3 and lanes number 4. And so notice that in lane number 3 and lane number 4 over here, that we have the plus inhibitor on top of it, indicating that the inhibitor described here, is utilized in lanes 3-4, but it is not utilized in lanes 1-2. And so it tells us that the results are shown down below and what conclusion can be made from the results or data. And so notice that the expected beta arrestin-bound GPCR band is right here in the middle. And so the only lane that has this expected beta arrestin-bound GPCR band is lane number 2. All of the other lanes do not have beta arrestin bound to the GPCR. And so essentially what we need to do is take a look at the conditions in lane number 2, and what we can see is that exposure to epinephrine is important in order for beta arrestin to bind. And so, that is definitely an important feature because when there's 0 minutes of exposure here in lane number 1, notice that there is no band under the same exact conditions except with 0 minutes of exposure. And then notice that, even if you do have exposure to epinephrine, if this inhibitor is present, then you will not have that band, you will not have beta arrestin bound to the GPCR. And so, this band right here in lane number 2 is telling us that in order for beta arrestin to bind to the GPCR, the receptor must be exposed and bound to its ligand. And so, that is really the main conclusion that we can make from this problem. So we can go ahead and indicate that a here is correct. Now taking a look at the option b, it says in order for beta arrestin to bind, the receptor cannot be exposed to its ligand. But of course when we take a look at the receptor not being exposed to any epinephrine, 0 minutes of exposure, notice that there is no band here. And so, of course this is not going to be the case because if beta arrestin bound, when there was no exposure, then we would expect to see a band here, but that's not what we see, so b is incorrect. And of course, c is also incorrect because it says in order for beta arrestin to bind, that BRC must first be bound to the receptor. But of course, we know that BRC does not bind to the receptor; it only phosphorylates the receptor since it is a kinase. And then of course d is also incorrect because it says in order for beta arrestin to bind, the inhibitor must first bind to the receptor. But of course, whenever the inhibitor is present, notice that the beta arrestin protein is never bound to the GPCR, which is why there are no bands here in lanes 3-4. And so again, a here is the correct answer to this example problem and that concludes this example so I'll see you guys in our next video.
How does GPCR termination differ from GPCR desensitization?
Which of the following statements about beta-arrestin and GPCRs is TRUE?
Which of the following are involved in the desensitization of the β-adrenergic receptor?
Here’s what students ask on this topic:
What is the difference between stimulatory and inhibitory adenylate cyclase GPCR signaling?
Stimulatory adenylate cyclase GPCR signaling involves the activation of adenylate cyclase by the stimulatory G protein (Gs), leading to increased production of the secondary messenger cAMP. In contrast, inhibitory adenylate cyclase GPCR signaling involves the inhibitory G protein (Gi), which inhibits adenylate cyclase activity, resulting in decreased cAMP levels. Both pathways regulate the activity of adenylate cyclase to ensure the appropriate cellular response to signals.
Created using AIHow does GPCR desensitization occur?
GPCR desensitization occurs when a signal is continuously present, leading to repeated exposure. This process involves the beta adrenergic GPCR kinase (BARK) phosphorylating the ligand-bound GPCR, followed by the binding of beta arrestin. Beta arrestin blocks the GPCR's interaction with the G protein and can trigger temporary endocytosis of the GPCR, making it inaccessible to the ligand. This dampens the cellular response despite the persistent presence of the primary messenger.
Created using AIWhat role does beta arrestin play in GPCR desensitization?
Beta arrestin plays a crucial role in GPCR desensitization by binding to the phosphorylated GPCR, which has been marked by beta adrenergic GPCR kinase (BARK). This binding prevents the GPCR from interacting with the G protein, thereby inhibiting the cellular response. Additionally, beta arrestin can initiate temporary endocytosis of the GPCR, removing it from the plasma membrane and further reducing its activity in response to the continuous ligand exposure.
Created using AIWhat is the function of the beta adrenergic GPCR kinase (BARK) in GPCR signaling?
The beta adrenergic GPCR kinase (BARK) is a kinase that phosphorylates the beta adrenergic GPCR when it is bound by a ligand. This phosphorylation marks the GPCR for binding by beta arrestin, which then blocks the GPCR's interaction with the G protein. This process is essential for GPCR desensitization, allowing the cell to dampen its response to continuous ligand exposure and preventing overstimulation.
Created using AIHow do stimulatory and inhibitory GPCR pathways regulate adenylate cyclase activity?
Stimulatory and inhibitory GPCR pathways regulate adenylate cyclase activity by acting like a gas pedal and brakes, respectively. The stimulatory pathway, mediated by the Gs protein, activates adenylate cyclase, increasing cAMP production. Conversely, the inhibitory pathway, mediated by the Gi protein, inhibits adenylate cyclase, decreasing cAMP levels. Together, these pathways ensure that adenylate cyclase produces the appropriate amount of cAMP needed for the cell's response to various signals.
Created using AI